
 PDF下载 ( 1147 KB)
PDF下载 ( 1147 KB)

-
摘要:
<正>【据《Indian J Gastroenterol》2013年4月报道】题:一项关于慢加急性肝衰竭病因、临床特点及在院病死率的前瞻性研究(作者Jha AK等)慢加急肝衰竭发生急性损伤的病因是多变的,有可能同时存在一种以上的急性损伤的因素。作者描述慢加急性肝衰竭的临床概况及自然病程中双重损伤因素的效果。
-
α/β干扰素共用Ⅰ型受体,该受体有α、β两个亚单位,分别称为IFNAR1和IFNAR2[1-4]。中国旱獭是研究HBV感染的重要动物模型[5-6]。本研究对中国旱獭Ⅰ型干扰素受体β亚基(marmata himalayana type Ⅰ interferon receptor β subunit,mhIFNAR2)进行了克隆、表达和抗体制备,并在体外用siRNA鉴定其功能,为研究干扰素抗土拨鼠肝炎病毒的确切机制、土拨鼠肝炎病毒持续感染的原因及提高干扰素的疗效奠定基础。
1. 材料与方法
1.1 材料
HepG2细胞、DH5α、BHK细胞、脑心肌炎病毒(encephalomyocarditis virus,EMCV)由同济医学院附属协和医院感染科实验室保存;WH12-6细胞由德国埃森大学陆蒙吉教授提供;pSUPER由美国Baylor大学细胞生物学系冯新华教授惠赠;引物和RNAi由上海英俊公司合成;限制性内切酶Pst Ⅰ、EcoR Ⅰ、Bgl Ⅱ和Sal Ⅰ、pMD18T载体、DL2000、T4 DNA连接酶、T4多核苷酸激酶购自Takara公司;Trizol、Lipofectamine2000TM购自Invitrogen公司;质粒抽提试剂盒为北京博大泰克生物基因技术有限公司生产;Taq酶和胶回收试剂盒为Tiangen公司产品;dNTP由上海凌飞公司提供;DMEM由HyClone公司提供;M-MLV逆转录酶、RNasin由美国Promega公司提供;小牛血清购自杭州四季青公司;凝胶成像分析系统购自美国UVP公司。根据其他哺乳动物干扰素受体的保守性设计引物,由上海英俊公司合成,其他哺乳动物在GenBank中的序列号分别为:鼠NM_010509,牛NM_174553,羊NM_001009342,人NM_207585(表1)。RT-PCR扩增目的基因mhIFNAR2(50-181aa)。
表 1 干扰素受体β亚基的引物序列Table 1. Primer sequences for interferon receptor β subunit名称 序列 核苷酸 位点 mhIFNAR1-2s 5′-AGTACATTTAGAAGCTGAAG-3' 396~416 mhIFNAR1-2as 5'-CTCTTCAGACCAAAAAGATG-3' 943~963 mhIFNAR2-1s 5'-CCATCTTATCATGGGAATTA-3' 149~169 mhIFNAR2-1as 5'-CTCTCAAAAACACAGAGTT-3' 1 281~1 300 1.2 方法
1.2.1 mhIFNAR2的克隆
根据其他哺乳动物干扰素受体的保守序列设计引物,克隆具体步骤见文献[7]。
1.2.2 Ⅰ型干扰素受体β亚基的表达及多克隆抗体的制备
测定重组质粒pRSET-B.mhIFNAR2核酸序列;重组质粒pRSET-B.mhIFNAR2转化高效表达菌Rossetta (DE3)plysS;菌体总蛋白的Western Blot分析:转膜后利用兔抗His-probe(H-15)作为一抗,HRP标记的羊抗兔IgG作为二抗进行Western Blot检测目的蛋白的表达;融合蛋白的大量制备及纯化:取纯化蛋白液20 μL,加2×蛋白电泳上样缓冲液20 μL于管中,100 ℃煮沸5 min。SDS-PAGE电泳,考马斯亮蓝染色观察纯化效果;检测蛋白纯度:将目的蛋白表达过程中的各个组分进行SDS-PAGE电泳,结果进行蛋白条带灰度扫描,并用Gelwork 1D advanced V4.01软件分析。检测蛋白浓度:通常每次纯化的样本都要行SDS-PAGE检测纯化样本的纯度,高纯度的样本被收集起来,用标准的Bradford法(考马斯亮蓝G-250法)测定蛋白浓度,为下一步免疫动物作准备。以纯化的重组蛋白mhIFNAR2B(50-181aa)为免疫原,每次取120 μg溶于1 mL PBS中,再与等体积的氢氧化铝佐剂混合,按40 μg/只的抗原剂量,分别免疫3只BALB/c小鼠,颈背部皮下多点注射。第14天和第28天时各加强免疫一次,加强免疫的方法和抗原剂量同上。第35天摘小鼠眼球取血,静置离心后分离血清。
1.2.3 多克隆抗体特异性鉴定
外周血单个核细胞 (peripheral blood mononuclear cell,PBMC)来源于中国旱獭,其分离的步骤如下:1 mL抗凝全血加入等体积的PBS混匀;将步骤1中的稀释全血用滴管沿管壁缓慢叠加在另一个装有2 mL的淋巴细胞分离液离心管内,注意保持清楚界面;2 500 r/min,离心20 min,管内液体分三层(从下至上为红色细胞层、分离液层、血浆层)。上层与中间层有一白色云雾状狭窄带,包括淋巴细胞、单核细胞、血小板;抽吸云雾层至灭菌管中,加入5倍体积以上的灭菌PBS(或D-Hank’s),2 000 r/min,离心5 min,弃上清;加入5倍体积以上的灭菌PBS(或D-Hank’s),2 000 r/min,离心5 min,弃上清;加入含10%小牛血清的RPMI1640 500 μL重悬细胞,每张APES处理的玻片上滴加100 μL,自然风干后用冰丙酮固定30 min行免疫组化及免疫荧光检测。用Western Blot检测其滴度。
1.2.4 mhIFNAR2功能的初步鉴定
1.2.4.1 中国旱獭α干扰素的制备
将生长状态良好的BHK细胞接种到6孔板内;生长24 h后,细胞长满至80%~90%;溶液A:2.5 μg质粒+Optim无血清培养基,总体积为100 μL;溶液B:5 μL Lipofectamine2000+95 μL Optim无血清培养基;两者室温放置5 min;混合溶液A和B,轻轻混匀,室温放置25 min;期间用无菌D-hank’s及培养基洗细胞两次,并加入1 600 μL无血清培养基;溶液A+溶液B内加入200 μL Optim无血清培养基,轻柔混匀后加入培养细胞孔内;37 ℃孵育5 h;换用完全培养基2.0 mL培养48 h后收获培养上清液,分装后冻存于-70 ℃,待测干扰素生物学活性。
1.2.4.2 EMCV感染滴度的测定
将生长在培养瓶内的WH12-6细胞消化后加入Ham’s-12培养基,轻轻吹打分散细胞后转种到96孔板内,100 μL/孔;待细胞在96孔板内长成单层后倾去培养液,滴定不同浓度的EMCV到细胞孔内,从原液逐渐至10-10,呈10倍系列稀释,100 μL/孔,同时做细胞阴性对照,每个梯度接种4孔细胞,第2天观察细胞生长状况,计数不同稀释度出现细胞病变效应(cytopathic effect,CPE)孔数,然后按Reed-Muench法计算半数组织培养感染剂量(TCID50)。
1.2.4.3 中国旱獭干扰素α效价的确定
将生长在培养瓶中的WH12-6细胞消化后加入Ham’s-12培养基,轻轻吹打分散细胞转种到96孔细胞培养板内;待细胞在96孔板内长成单层后倾去培养液,加入不同4倍系列稀释的BHK细胞转染上清100 μL/孔,每个稀释度有4个细胞孔,继续培养24 h,加入0.1 mL EMCV,继续培养24 h,以保护50%细胞不出现CPE的BHK细胞转染上清液最高稀释度作为干扰素效价。
1.2.4.4 siRNA干扰实验
WH12-6细胞培养于含10% FBS的Ham’s-12培养基中,于5% CO2、饱和湿度及37 ℃条件下培养。勿使细胞融合度>90%以防止细胞老化。siRNA由广州锐博公司提供合成,序列见表2。另外设计一条阴性对照序列,Blast验证为任何物种的无关序列。
表 2 mhIFNAR2 siRNA 模板序列Table 2. Template of mhIFNAR2 siRNAcDNA 正义链(5′-3′) 反义链(3′-5′) GAAGTAGCTCTCAGAACTA(mhIFNAR1B840B) GAAGUAGCUCUCAGAACUAdTdT dTdTCUUCAUCGAGAGUCUUGAU GCTTTACAGACCACATTAA(mhIFNAR2B277B) GCUUUACAGACCACAUUAAdTdT dTdTCGAAAUGUCUGGUGUAAUU GCTGAAGATAAGGCAATAA(mhIFNAR1B412B) GCUGAAGAUAAGGCAAUAAdTdT dTdTCGACUUCUAUUCCGUUAUU 阴性对照 公司未提供序列 将正常培养于6孔板的WH12-6细胞于转染前改用无抗生素Ham’s-12培养基培养24 h。取2 μL Lipofectamine 2000加入250 μL Opti-MEM培养基,轻轻混匀。室温孵育5 min。取20 μmol/L合成siRNA,加入250 μL Opti-MEM培养基,siRNA的终浓度分10、20及50 nmol/L三个梯度,轻轻混匀,室温孵育5 min。随后将两者轻轻混合,室温孵育20 min。加入一孔细胞中,用无血清无抗生素Ham’s-12培养基定容至2 mL。于37 ℃、5% CO2培养箱中培养6 h。换为含血清、抗生素的全培养基,培养24 h后加用中国旱獭干扰素刺激(同前),24 h后加Trizol提取RNA。用Trizol提取培养的WH12-6细胞总RNA,逆转录成cDNA,使用SYBR Green嵌合荧光法进行RT-PCR扩增,反应条件为95 ℃、60 s预变性,95 ℃、15 s共40个循环,60 ℃、20 s延伸。mhIFNAR2、MxA荧光定量RT-PCR引物序列如下:mhIFNAR2(正义)ACAATCCACCGTGCCAACT,mhIFNAR2(反义)AACAGCATCGCTTTCCCTC;MxA(正义)CAGATTGTCTACTGCCAGGACCAGGT,MxA(反义)AGCCGCTCCTTCAGGAACTTC;β-actin(正义)TGGAATCCTGTGGCATCCATGAAAC,β-actin(反义)TAAAACGCAGCTCAGTAACAGTCCG;通过标准曲线获得直线回归方程,从而得到各个目的基因mRNA的拷贝数,将其除以内参的拷贝数,然后乘以104作为该目的基因的相对值。
1.3 统计学方法
采用SPSS 18.0软件进行统计学分析。计量资料以
2. 结果
2.1 mhIFNAR2的克隆、鉴定及同源性比较
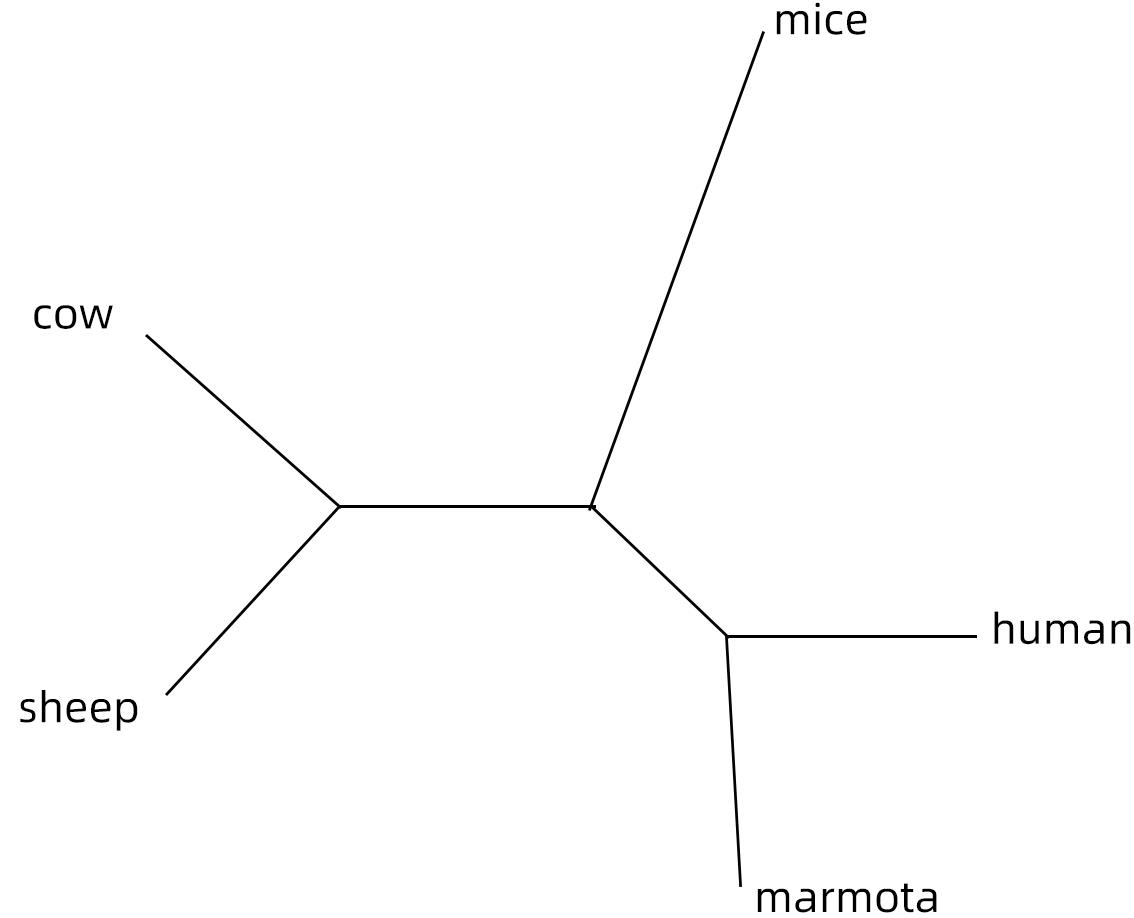
RT-PCR从脾组织中扩增出149~1 300 bp的mhIFNAR2片段,并经测序证实[7]。同源重组构建重组质粒,经PCR、酶切和测序证实,命名为pRSET-B.mhIFNAR2。mhIFNAR2149-1300核苷酸序列与土拨鼠、人、牛、羊及小鼠的同源性分别为98.05%、72.89%、69.17%、69.76%及64.95%(图1)。
2.2 重组蛋白的表达
2.2.1 重组质粒核苷酸组成示意图 ATG---6×His---Pst ImhIFNAR250-181 TGA---B BEcoR I
2.2.2 重组蛋白的小量诱导表达
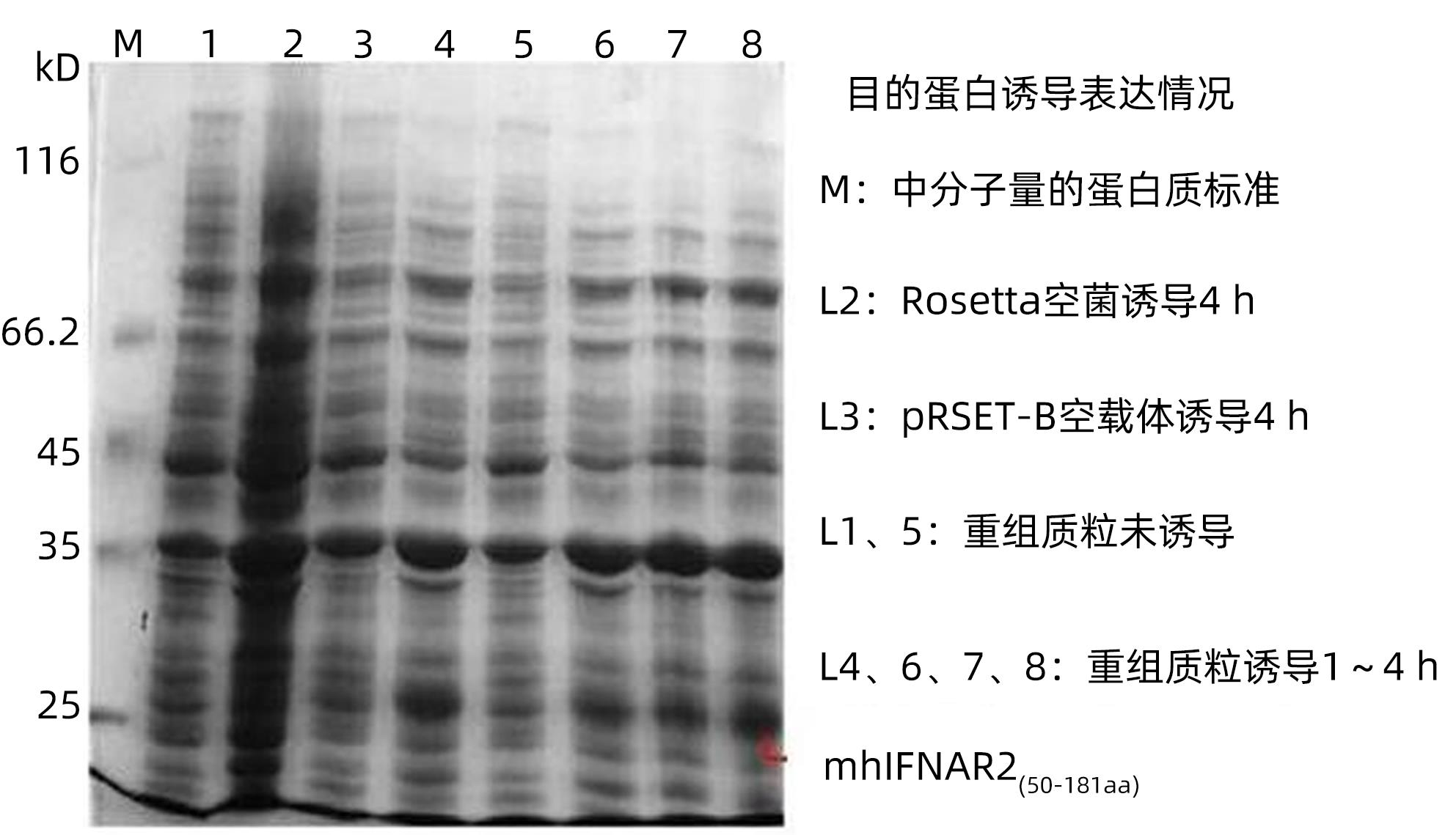
SDS-PAGE电泳可见,pRSET-B.mhIFNAR2重组质粒经IPTG诱导后1~6 h在大肠杆菌中均有表达,且在诱导4 h时获得量最大,而空菌及空载体转化菌诱导4 h在27 kD处仅有较弱条带(图2)。
 图 2 SDS-PAGE电泳分析His-mhIFNAR2蛋白的表达Figure 2. Expression of His-mhIFNAR2 analyzed by SDS-PAGE
图 2 SDS-PAGE电泳分析His-mhIFNAR2蛋白的表达Figure 2. Expression of His-mhIFNAR2 analyzed by SDS-PAGE2.2.3 目的蛋白的纯化
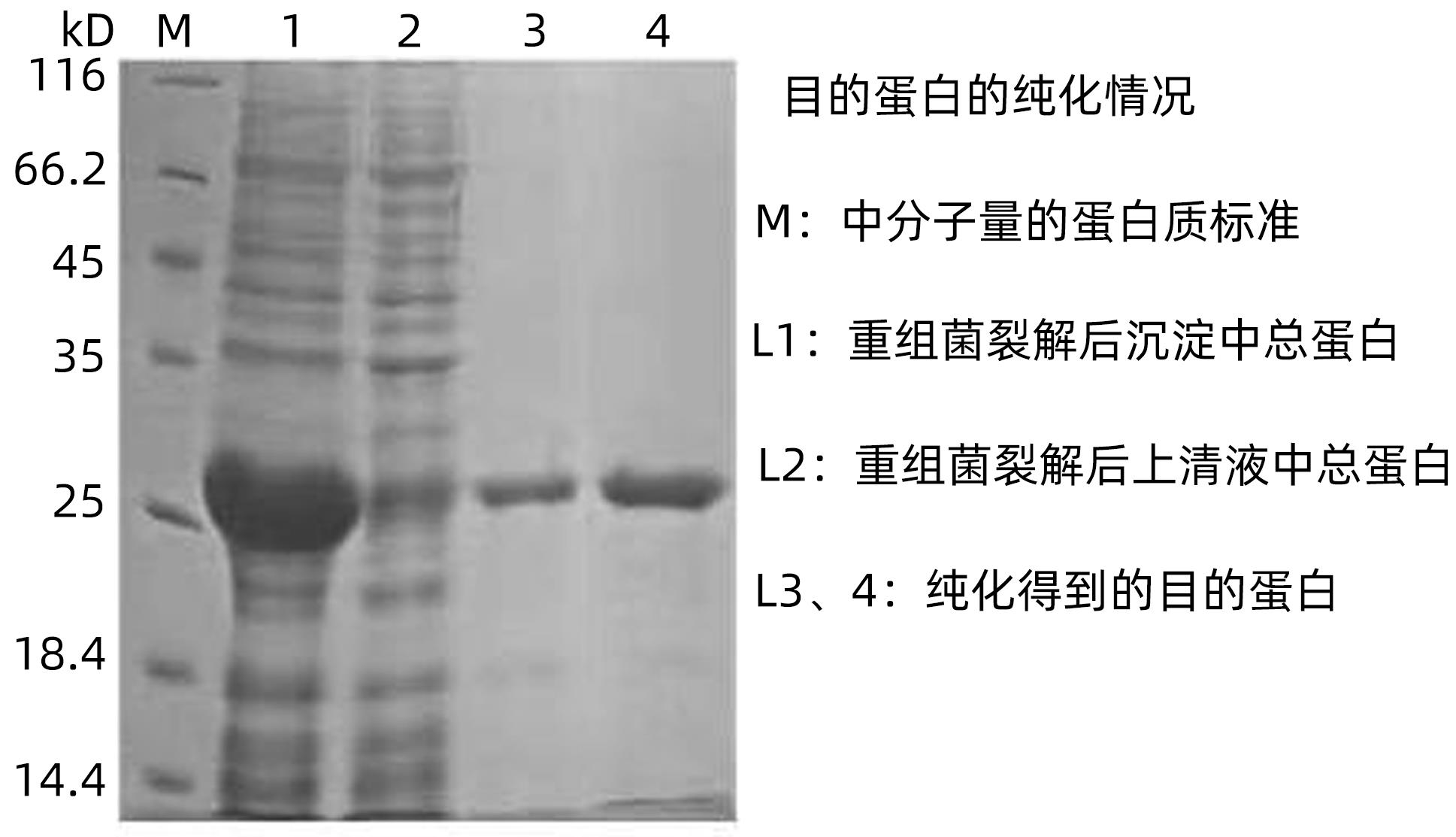
诱导后的菌体超速离心后,经SDS-PAGE电泳发现目的蛋白大部分存在于沉淀中,且主要以包涵体的形式存在。经Ni-NTA纯化得到的目的蛋白纯度为95%,浓度约为160 μg/mL(图3)。
2.2.4 目的蛋白的Western Blot检测
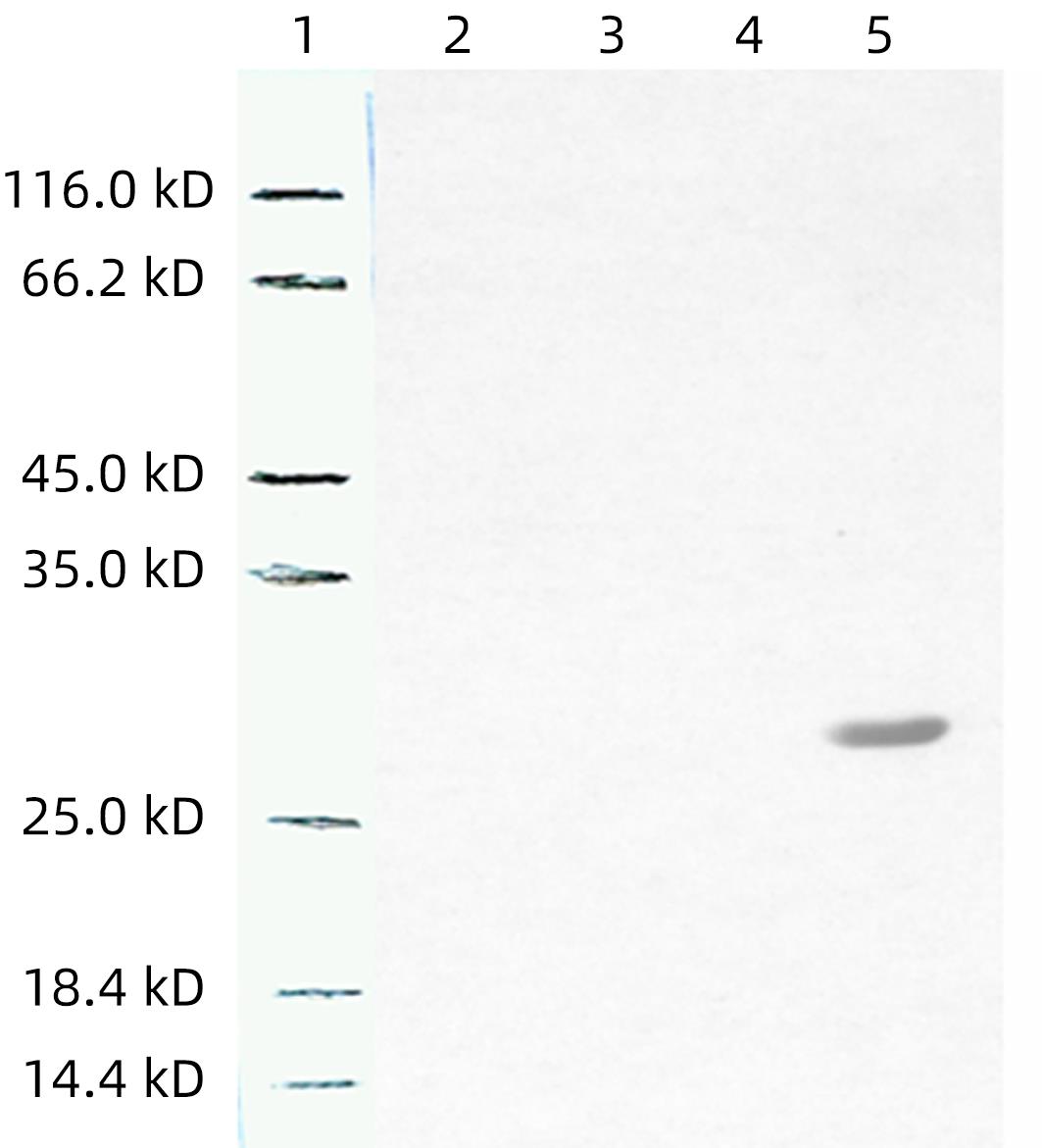
抗His与纯化的重组蛋白反应,可见有特异性条带(图4)。
 注: L1,中分子量的蛋白质标准;L2,mhIFNAR2多抗血清作为一抗(1∶1 000);L3,mhIFNAR2多抗血清作为一抗(1∶10 000);L4,正常小鼠血清作为一抗;L5,兔抗His作为一抗。图 4 纯化的目的蛋白Western Blot检测Figure 4. Western Blot analysis of the activity and specificity of antisera
注: L1,中分子量的蛋白质标准;L2,mhIFNAR2多抗血清作为一抗(1∶1 000);L3,mhIFNAR2多抗血清作为一抗(1∶10 000);L4,正常小鼠血清作为一抗;L5,兔抗His作为一抗。图 4 纯化的目的蛋白Western Blot检测Figure 4. Western Blot analysis of the activity and specificity of antisera2.3 制备抗体的检测
2.3.1 免疫组化检测抗mhIFNAR2多克隆抗体特异性
抗原抗体结合部位可见明显的阳性着色(图5a),mhIFNAR2抗原的分布主要呈胞质型和胞膜型。而应用正常小鼠血清作为一抗则无阳性显色(图5b)。
 图 5 mhIFNAR2多克隆抗体的免疫组化结果(DAB,×400)Figure 5. Immunohistochemical of mhIFNAR2 polyclonal antibodies(DAB,×400)
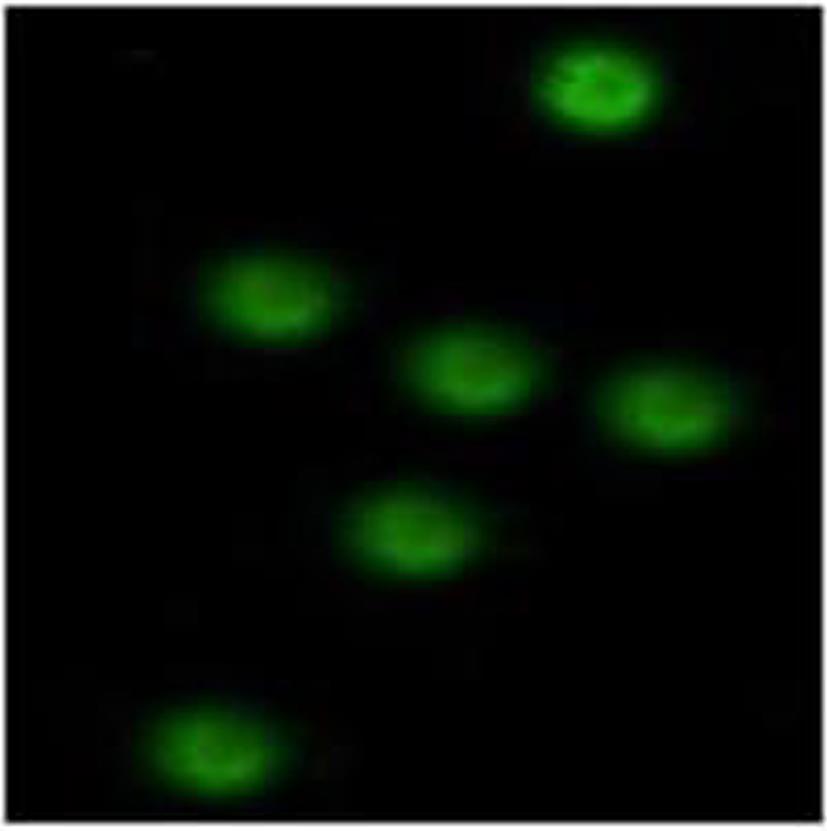
图 5 mhIFNAR2多克隆抗体的免疫组化结果(DAB,×400)Figure 5. Immunohistochemical of mhIFNAR2 polyclonal antibodies(DAB,×400)2.3.2 免疫荧光结果
PBMC中mhIFNAR2免疫荧光检测主要分布于细胞质和细胞膜(图6),而阴性对照则无阳性显色。
 图 6 PBMC中mhIFNAR2免疫荧光检测Figure 6. Stained with mouse anti-mhIFNAR2 antibody by immunofluorescence in PBMC
图 6 PBMC中mhIFNAR2免疫荧光检测Figure 6. Stained with mouse anti-mhIFNAR2 antibody by immunofluorescence in PBMC2.3.3 Western Blot检测
用纯化的重组蛋白免疫BALB/c小鼠后,获得了高效价的特异性多克隆抗体。经用Western Blot检测的效价为1∶1 000。
2.4 干扰素效价的测定
以保护50%的细胞不出现CPE的BHK细胞转染上清稀释度作为干扰素的效价,表3为EMCV感染滴度测定结果,本实验所测得的干扰素效价为1∶128。
表 3 EMCV感染滴度测定结果Table 3. Result of EMCV infection titer determination病毒稀释 有CPE孔 无CPE孔 累计有CPE孔 累计无CPE孔 有/总和 感染百分率(%) 100 4 0 39 0 39/39 100 10-1 4 0 35 0 35/35 100 10-2 4 0 31 0 31/31 100 10-3 4 0 27 0 27/27 100 10-4 4 0 23 0 23/23 100 10-5 4 0 19 0 19/19 100 10-6 4 0 15 0 15/15 100 10-7 4 0 11 0 11/11 100 10-8 4 0 7 0 7/7 100 10-9 2 2 3 2 3/5 60 10-10 1 3 1 5 1/6 16.7 2.5 siRNA干扰效应
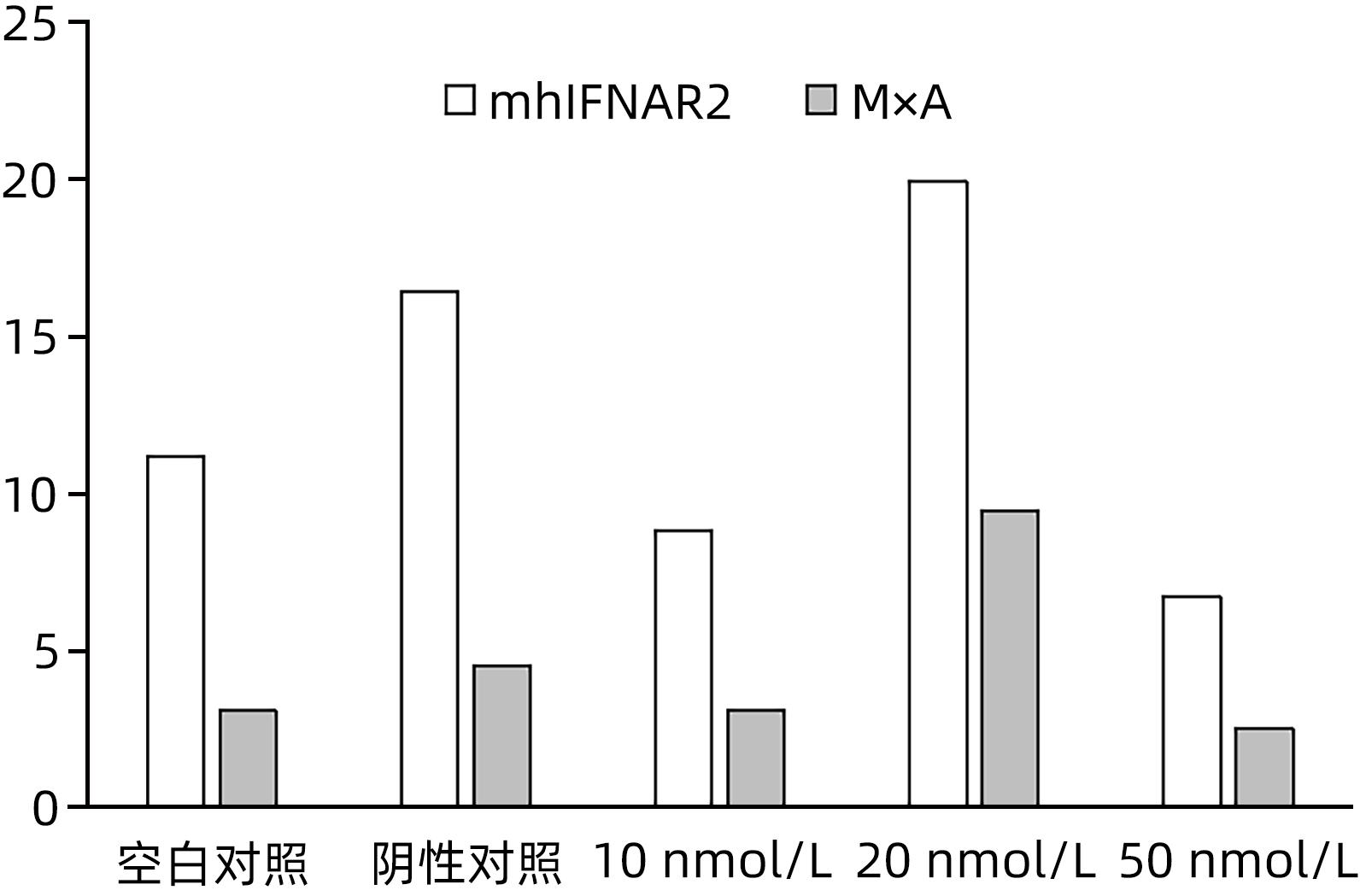
起始于840位点的siRNA840对WH12-6细胞mhIFNAR2基因无明显的抑制作用(P>0.05)(图7)。起始于277位点的siRNA277(20 nmol/L)对WH12-6细胞mhIFNAR2、M×A基因有明显抑制作用,差异均具有统计学意义(P值均<0.05)(表4)。
 图 7 不同浓度的siRNA840对WH12-6细胞mhIFNAR2、M×A基因的抑制作用Figure 7. The inhibitory effect of siRNA840 at different concentrations on mhIFNAR2 and M×A表 4 不同浓度的siRNA277对WH12-6细胞mhIFNAR2、M×A基因的抑制作用Table 4. The inhibitory effect of siRNA277 at different concentrations on mhIFNAR2 and M×A
图 7 不同浓度的siRNA840对WH12-6细胞mhIFNAR2、M×A基因的抑制作用Figure 7. The inhibitory effect of siRNA840 at different concentrations on mhIFNAR2 and M×A表 4 不同浓度的siRNA277对WH12-6细胞mhIFNAR2、M×A基因的抑制作用Table 4. The inhibitory effect of siRNA277 at different concentrations on mhIFNAR2 and M×A组别 mhIFNAR2 M×A 空白对照(n=3) 11.678±0.725 3.087±1.910 阴性对照(n=3) 14.190±3.100 4.150±0.500 20 nmol/L(n=3) 2.538±0.1241)2) 1.350±0.2121)2) 注:与空白对照组比较,1)P<0.05;与阴性对照组比较,(2)P<0.05。 3. 讨论
干扰素受体由α、β两个亚基组成,当其与干扰素结合后迅速引起TYK2、JAK1、STAT1、STAT2磷酸化,进而激活转录产生各种抗病毒蛋白。尽管干扰素发现已有50年的历史,但直到1990年才鉴定出干扰素受体由α、β两个亚基组成。其中β亚基有长、短及可溶等三种形式[8-10]。随着干扰素与其受体相互作用、信号转导机制及生物应答研究的深入,干扰素受体在介导干扰素抗感染中的天然免疫、获得性免疫以及肿瘤中的作用逐渐引起人们的注意,干扰素受体缺失引起严重的病原体感染[11-12]。但是,干扰素在不同的细胞中会产生相反的生物学效应。比如在大多数细胞中干扰素表现为抑制增殖,促进凋亡,却可延长记忆性T淋巴细胞的存活时间[13-14]。因此,阐明干扰素受体复合物的功能有助于解释这些现象的原因。在乙型肝炎动物模型中阐明干扰素抗病毒的作用机制及HBV的持续性感染具有重要的临床意义。
本研究利用哺乳动物干扰素受体序列的保守区设计引物,成功地从中国旱獭脾组织中克隆出mhIFNAR2149-1300。mhIFNAR2149-1300核苷酸序列与土拨鼠、人、牛、羊及小鼠的同源性分别为98.05%、72.89%、69.17%、69.76%及64.95%。该结果与笔者前期克隆的中国旱獭其他分子相似[15-18]。
干扰素是具有多种生物功能的细胞因子,其发挥效应的始动环节是与其受体结合,使胞内段发生磷酸化,激活转录从而产生各种抗病毒蛋白。本研究目的是在mhIFNAR2克隆的基础上,在体外用抗体阻断及siRNA干扰的方法进一步证实所克隆基因的正确性,了解mhIFNAR2在干扰素信号通路的作用,为在乙型肝炎动物模型中国旱獭体内实验奠定基础。
前期研究[14]结果显示IFNAR2与干扰素结合的重要位点包括E78、W101、I104以及D105,而既往的一些研究[19-20]也发现干扰素受体胞外段作为免疫原可以产生中和抗体。因此本文选择了mhIFNAR2(50-181aa)胞外段重组蛋白作为抗原制备抗体。
本实验以胞外段mhIFNAR2重组蛋白免疫小鼠,制备了抗mhIFNAR2的多克隆抗体。对所获得的抗血清进行了Western Blot检测,证实制备的多克隆抗体特异性良好,具有免疫学活性,不仅可以识别作为免疫原的融合蛋白,还能有效识别PBMC中的mhIFNAR2蛋白。免疫组化及免疫荧光检测可见mhIFNAR2抗原的分布主要呈胞浆型和胞膜型。并且免疫血清在较高稀释浓度下即可检测到mhIFNAR2蛋白,且无明显的非特异性条带,证实了多克隆抗体的高效性和抗mhIFNAR2的特异性,为进一步研究mhIFNAR2及干扰素在乙型肝炎动物模型中的作用提供了实验基础。
本研究用该抗体进行干扰素受体封闭实验,结果显示无明显作用(具体数据未展示),原因可能有:(1)某些抗体虽然可以与其受体结合,但是不能阻断干扰素的作用,且不同干扰素型别存在差异[21-22];(2)不同细胞系与抗体的结合力也存在差异[17];(3)一些抗体并不能阻止干扰素与其受体结合[22]。此外,本研究制备的是多克隆抗体,一是没有纯化,二是某些多克隆位点可能对干扰素信号通路产生影响。
本研究设计、合成针对中国旱獭mhIFNAR2的siRNA,并在基因水平成功验证了起始于277位点的siRNA能够有效抑制mhIFNAR2的表达,为深入研究mhIFNAR2的功能和机制,尤其是为在乙型肝炎中的研究提供了一个手段。
-

 下载:
下载:


 下载:
下载:

