
 PDF下载 ( 5284 KB)
PDF下载 ( 5284 KB)

细胞周期素D1对HBV转录和复制的影响
DOI: 10.3969/j.issn.1001-5256.2023.02.010
-
摘要:
目的 探究细胞周期素D1(cyclin D1,基因名CCND1)对HBV复制的影响及其机制。 方法 利用GSE84044数据集,采用Spearman秩相关分析HBV相关肝纤维化患者肝组织基因表达水平与血清HBV DNA载量之间的相关性。在HBV细胞复制模型中瞬时表达cyclin D1及cyclin D1持续激活突变体(T286A)蛋白,使用时间分辨免疫荧光及实时荧光定量PCR实验分别检测细胞培养上清液中的HBsAg、HBeAg及HBV DNA水平,Western blot检测细胞内HBV core蛋白,反转录-实时荧光定量PCR法检测细胞内HBV RNA,双荧光素酶报告基因实验检测cyclin D1对HBV基本核心启动子(BCP)活性的影响。利用GSE83148数据集,分析CCND1与HBV相关调控因子表达的相关性。正态分布的计量资料两组间比较采用独立样本t检验;非正态分布的计量资料两组间比较采用Mann-Whitney U检验。 结果 在GSE84044数据中,HBV相关肝纤维化患者的肝组织中有7个细胞周期调控基因与HBV DNA载量呈显著负相关(r值均<-0.3,P值均<0.05),其中包括CCND1基因(r=-0.474,P<0.001)。外源表达cyclin D1及cyclin D1 T286A突变体能降低HBV复制细胞模型培养上清液中的HBsAg、HBeAg及HBV DNA水平,以及细胞内core蛋白和HBV RNA水平;外源表达cyclin D1显著抑制了HBV BCP的转录活性;且慢性乙型肝炎患者肝组织中CCND1表达水平与抑制HBV复制的APOBEC3G(r=0.575,<0.001)、SMC5(r=0.341,P<0.001)和FOXM1(r=0.333,P<0.001)表达呈显著正相关,而与HBV进入受体NTCP(r=-0.511,P<0.001)和HBV复制正向调控转录因子HNF1α(r=-0.430,P<0.001)表达呈显著负相关。在HepG2细胞中过表达cyclin D1降低HNF1α及NTCP转录水平。 结论 cyclin D1抑制HBV的转录和复制,可能与其下调HNF1α及NTCP表达有关。 Abstract:Objective To investigate the effect of cyclin D1 (with CCND1 as the gene name) on HBV replication and its potential mechanism. Methods With reference to GSE84044 dataset, the Spearman's rank correlation analysis was used to investigate the correlation between the expression levels of genes in liver tissue and serum HBV DNA load in patients with HBV-related liver fibrosis. Cyclin D1 and cyclin D1 T286A mutant were transiently expressed in the HBV cell replication model, and time-resolved immunofluorescence and quantitative real-time PCR were used to measure the levels of HBsAg/HBeAg and HBV DNA in cell culture supernatant; Western blot was used to measure the level of HBV core protein in cells; reverse-transcription quantitative real-time PCR was used to measure the level of HBV RNA in cells; dual-luciferase reporter assay was used to observe the effect of cyclin D1 on the activity of HBV basic core promoter (BCP). GSE83148 dataset was used to investigate the correlation between CCND1 and HBV-related regulatory factors. The independent samples t-test was used for comparison of normally distributed continuous data between two groups, and the Mann-Whitney U test was used for comparison of non-normally distributed continuous data between two groups. Results The analysis of GSE84044 data showed that 7 cell cycle genes were significantly negatively correlated with HBV DNA load in liver tissue of the patients with HBV-related liver fibrosis (all r < -0.3, all P < 0.05), which included the CCND1 gene (r=-0.474, P < 0.001). Exogenous expression of cyclin D1 and cyclin D1 T286A mutant reduced the levels of HBsAg, HBeAg, and HBV DNA in culture supernatant of the HBV replication cell model, as well as the levels of HBV core protein and HBV RNA in cells. Exogenous expression of cyclin D1 significantly inhibited the transcriptional activity of HBV BCP. The expression level of CCND1 in liver tissue of chronic hepatitis B patients was significantly positively correlated with the expression of APOBEC3G (r=0.575, P < 0.001), SMC5 (r=0.341, P < 0.001), and FOXM1 (r=0.333, P < 0.001) which inhibited HBV replication, while it was significantly negatively correlated with the expression of the HBV entry receptor NTCP (r=-0.511, P < 0.001) and HNF1α as the transcription factor for positive regulation of HBV replication (r=-0.430, P < 0.001). Overexpression of cyclin D1 in HepG2 cells reduced the transcriptional levels of HNF1α and NTCP. Conclusion Cyclin D1 inhibits HBV transcription and replication possibly by downregulating the expression of HNF1α and NTCP. -
Key words:
- Hepatitis B Virus /
- Cyclin D1 /
- Virus Replication /
- Gene Transcription
-
HBV感染是全球重要公共卫生问题之一[1-2]。但由于对HBV生命周期的认识还不够充分,目前尚缺乏有效治愈慢性乙型肝炎(CHB)的药物。因此,深入解析HBV复制的调控机制,将为抗HBV新药物研发提供有效靶点。
HBV属嗜肝DNA病毒科病毒,基因组为部分双链的松弛环状DNA(relaxed circular DNA, rcDNA),其复制过程有着独特的逆转录步骤[3]。研究[4-5]表明,HBV的复制依赖于肝细胞的细胞周期。例如,HBV在静止肝细胞中的复制更为活跃,而在细胞开始分裂时复制减慢。细胞周期素D1(cyclin D1,基因名CCND1)是细胞周期进程的正性调节因子,可通过结合并激活细胞周期素依赖性激酶(cyclin-dependent kinase, CDK)4、6,促进细胞由静止期(G0)或间期1(G1)进入DNA合成期(S)。研究[6-7]发现,cyclin D1在包括肝癌在内的多种肿瘤组织中呈异常高表达,但cyclin D1是否影响HBV复制目前尚未见报道。本研究通过挖掘公共数据库数据以及应用HBV复制细胞模型,探究cyclin D1对HBV复制的影响,并对其调控HBV复制的可能机制进行分析。
1. 材料与方法
1.1 材料
人肝癌细胞系HepG2购自美国ATCC数据库,Huh-7细胞购自上海中科院细胞库。pFLEX-cyclin D1-T286A、pGL3-HBV BCP、Actin-Renilla和pGL3-basic质粒为本室前期保存构建。prcccDNA/pCMV-Cre重组质粒系统由prcccDNA质粒和pCMV-Cre表达质粒组成[8],为复旦大学基础医学院邓强教授惠赠。
1.2 细胞培养与转染
HepG2和Huh-7细胞均采用含10%胎牛血清和100 U/mL青霉素、链霉素的DMEM培养基,在37 ℃和含5% CO2的细胞培养箱中常规培养。细胞转染前1天,将生长状态良好的细胞按1.5×105个/孔接种至12孔板,其中HepG2细胞接种前需铺好鼠尾胶原。按照LipofectamineTM 2000(购自美国Invitrogen公司)说明书进行质粒转染,6 h后更换新鲜培养基继续培养,72 h后收集培养上清及细胞沉淀用于后续实验研究。
1.3 质粒构建
采用同源重组的方法构建pCMV-cyclin D1表达质粒,其中扩增cyclin D1编码区的引物序列为:上游5′-ATTGTACCCGCGGGCCATGGAACACCAGCTCCTGTGC-3′,下游5′-GGATCCCCGCGGCCGCGTCACTTGTCATCGTCATCCTTGTAATC-3′。cyclin D1持续激活突变体表达质粒pCMV-cyclin D1-T286A是在pFLEX-cyclin D1-T286A质粒基础上,采用双酶切连接的方法构建而成。上述质粒DNA的序列均经测序验证[测序由生工生物工程(上海)股份有限公司完成]。
1.4 HBsAg和HBeAg检测
使用时间分辨免疫荧光法检测细胞培养上清中HBsAg和HBeAg,检测试剂盒购自苏州新波生物技术公司。所用检测设备为SYM-BIO Anytest时间分辨荧光分析仪(上海新波生物技术有限公司)。
1.5 HBV DNA定量检测
细胞培养上清用3000 r/min离心3 min,吸取200 μL上清首先进行95 ℃加热10 min,使病毒蛋白变性,然后使用Light Cycle 480Ⅱ实时荧光定量PCR仪和罗氏2×SYBR Green Mix进行HBV DNA相对定量检测。上游引物序列为5′-CGGCGTTTTATCATMTTCCTCT-3′,下游引物序列为5′-GACAAACGGGCAACATACCTT-3′。
1.6 反转录-实时荧光定量PCR(RT-qPCR)实验
Trizol法提取细胞总RNA,采用Roche反转录试剂盒(购自美国罗氏公司)逆转录为cDNA。取2 μL cDNA为模板,使用2×SYBR Green Mix(购自美国罗氏公司)进行实时荧光定量PCR检测细胞内HBV mRNA表达水平,以管家基因ACTIN作为内参。HBV RNA检测所用引物序列如下:3.5 kb HBV RNA上游引物为5′-AGACCACCAAATGCCCCTATC-3′,下游引物为5′-TCTGCGAGGCGAGGGAGTTC-3′,可检测pgRNA和preC mRNA两种RNA;preC mRNA上游引物为5′-TCTGCGCACCAGCACCATG-3′,下游引物为5′-CAATGCTCAGGAGACTCTAAGGC-3′。宿主基因APOBEC3G上游引物为5′-GCATCGTGACCAGGAGTATGA-3′,下游引物为5′- GTCAGGGTAACCTTCGGGT-3′;牛磺胆酸钠共转运多肽(sodium taurocholate cotransporting polypeptide, NTCP)上游引物为5′-AAGGACAAGGTGCCCTATAAAGG-3′,下游引物为5′-TTGAGGACGATCCCTATGGTG-3′;肝细胞核因子1α(hepatocyte nuclear factor 1α, HNF1α)上游引物为5′-AACACCTCAACAAGGGCACTC-3′,下游引物为5′-CCCCACTTGAAACGGTTCCT-3′;染色体结构维持复合物5(structural maintenance of chromosome 5, SMC5)上游引物为5′-TCCCGAGAGACCCTTCGTC-3′,下游引物为5′-TTCCATTGGCTCCAACGATCA-3′;叉头盒转录基因M1(forkhead box M1, FOXM1)上游引物为5′-ATACGTGGATTGAGGACCACT-3′,下游引物为5′-TCCAATGTCAAGTAGCGGTTG-3′;ACTIN的上游引物为5′-TTGTTACAGGAAGTCCCTTGCC-3′,下游引物为5′-ATGCTATCACCTCCCCTGTGTG-3′。
1.7 Western blot实验
收集细胞沉淀,加入约细胞体积3倍的RIPA裂解液,冰上裂解30 min,12 000×g、4 ℃离心10 min后取上清,BCA蛋白定量试剂盒测定蛋白浓度。SDS-PAGE电泳分离蛋白后转移至PVDF膜上,5%脱脂奶粉室温封闭60 min,按各抗体说明书使用量稀释后加入适量一抗4 ℃摇床杂交过夜,加入二抗(二抗原液按1∶8000稀释)室温孵育2 h。使用Odyssey双色红外激光成像系统(美国LICOR公司)和Odyssey V1.2软件进行杂交膜的扫描和分析。
1.8 双荧光素酶活性检测
为了确定cyclin D1对HBV基本核心启动子(BCP)活性的影响,将带有HBV BCP区的pGL3荧光素酶报告载体(pGL3-HBV BCP)与pCMV-cyclin D1或pCMV-cyclin D1-T286A共转染HepG2细胞,同时共转Actin-Renilla质粒作为内参对照。转染48 h后,使用Dual Luciferase Reporter Assay试剂盒中的Passive Lysis Buffer裂解细胞,吸取25 μL上清检测萤火虫和海肾萤光素酶活性,以萤火虫/海肾的比值表示报告基因的活性。所用设备为多功能酶标仪(PerkinElmer公司)。
1.9 数据下载与分析
在GEO数据库(Gene Expression Omnibus,
https://www.ncbi.nlm.nih.gov/geo )下载转录组数据集GSE84044和GSE83148[9-10]。GSE84044收录了124例HBV相关肝纤维化患者肝组织的转录组数据,GSE83148收录了122例CHB患者肝组织的转录组数据。通过limma包[11],对下载的数据进行了归一化处理。在分析与HBV DNA相关的宿主基因时,选择了GSE84044数据集中性别为男性且血清HBV DNA载量大于104拷贝/mL的患者共计59例。1.10 统计学方法
采用GraphPad Prism V5.0、R(v4.1.3)[12]及SPSS 19.0统计软件进行数据分析。正态分布的计量资料两组间比较采用独立样本t检验;非正态分布的计量资料两组间比较采用Mann-Whitney U检验。相关性分析时采用Spearman秩相关,错误发现率(false discovery rate, FDR)<0.05且|r|>0.3被定义为显著相关基因。P<0.05为差异有统计学意义。
2. 结果
2.1 肝组织CCND1表达水平与血清HBV DNA载量呈负相关
通过分析GSE84044数据集中59例男性HBV相关肝纤维化患者肝组织的转录组数据,共找到了765个表达水平与患者血清HBV DNA水平显著相关的宿主基因(FDR<0.05,|r|>0.3,图 1a)。通过与116个细胞周期基因(ko04110)取交集,发现其中有7个基因为细胞周期相关基因,且这7个基因在肝脏中的表达水平均与血清HBV DNA水平呈显著负相关(FDR<0.05,r值均<-0.3,P值均<0.05) (图 1a、1b),按相关性由强到弱排序依次为MYC、TGFB1、CCND1、YWHAZ、HDAC1、YWHAB和E2F3。其中,CCND1是调控细胞从G0/G1期向S期转换的关键因子,其表达水平与HBV DNA载量呈显著负相关(r=-0.474,P<0.001)(图 1c)。
 图 1 肝组织基因表达与血清HBV DNA的相关性分析注:a,GSE84044数据集中与患者HBV DNA载量相关的肝组织宿主基因,样本按照HBV DNA载量的高低从左到右排列,蓝色代表低表达,红色代表高表达;b,765个与HBV DNA载量显著相关基因和116个细胞周期基因的交集;c,肝组织中CCND1水平与血清HBV DNA载量之间的相关性分析。Figure 1. Correlation analysis of host gene expression in liver tissue and serum HBV DNA level
图 1 肝组织基因表达与血清HBV DNA的相关性分析注:a,GSE84044数据集中与患者HBV DNA载量相关的肝组织宿主基因,样本按照HBV DNA载量的高低从左到右排列,蓝色代表低表达,红色代表高表达;b,765个与HBV DNA载量显著相关基因和116个细胞周期基因的交集;c,肝组织中CCND1水平与血清HBV DNA载量之间的相关性分析。Figure 1. Correlation analysis of host gene expression in liver tissue and serum HBV DNA level2.2 外源表达cyclin D1抑制HBV复制
为探究cyclin D1蛋白对HBV复制的影响,构建了pCMV-cyclin D1和pCMV-cyclin D1-T286A表达质粒,并将上述质粒分别与prcccDNA/pCMV-Cre质粒体系共同转染至HepG2及Huh-7细胞中,72 h后收集细胞沉淀和细胞培养上清液。Western blot实验证实pCMV-cyclin D1和pCMV-cyclin D1-T286A表达质粒均可在两种细胞中成功表达,且cyclin D1和cyclin D1(T286A)过表达均能显著降低细胞内HBV core蛋白水平(图 2a、2b)。与此结果一致,过表达cyclin D1和cyclin D1(T286A)亦显著下调了两种细胞上清液中的HBsAg、HBeAg和HBV DNA水平(图 2c、d)。
 图 2 外源表达cyclin D1降低HBV的复制水平注:a、b,外源表达cyclin D1降低HepG2和Huh-7细胞HBV core蛋白的水平;c、d,外源表达cyclin D1下调HepG2和Huh-7细胞培养上清液中的HBsAg、HBeAg与HBV DNA水平。Figure 2. Exogenous expression of cyclin D1 reduces the replication level of HBV
图 2 外源表达cyclin D1降低HBV的复制水平注:a、b,外源表达cyclin D1降低HepG2和Huh-7细胞HBV core蛋白的水平;c、d,外源表达cyclin D1下调HepG2和Huh-7细胞培养上清液中的HBsAg、HBeAg与HBV DNA水平。Figure 2. Exogenous expression of cyclin D1 reduces the replication level of HBV2.3 外源表达cyclin D1在转录水平抑制HBV复制
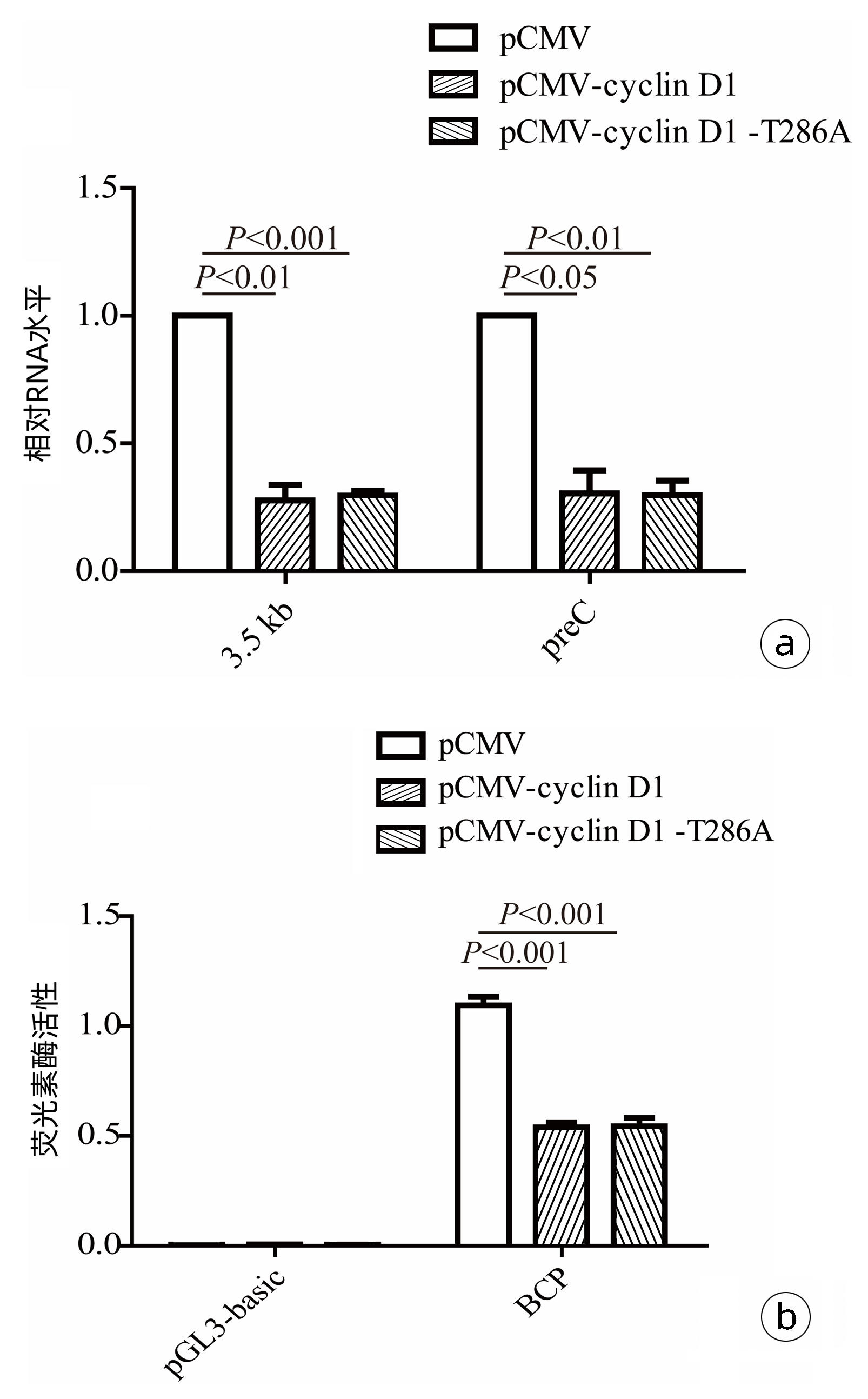
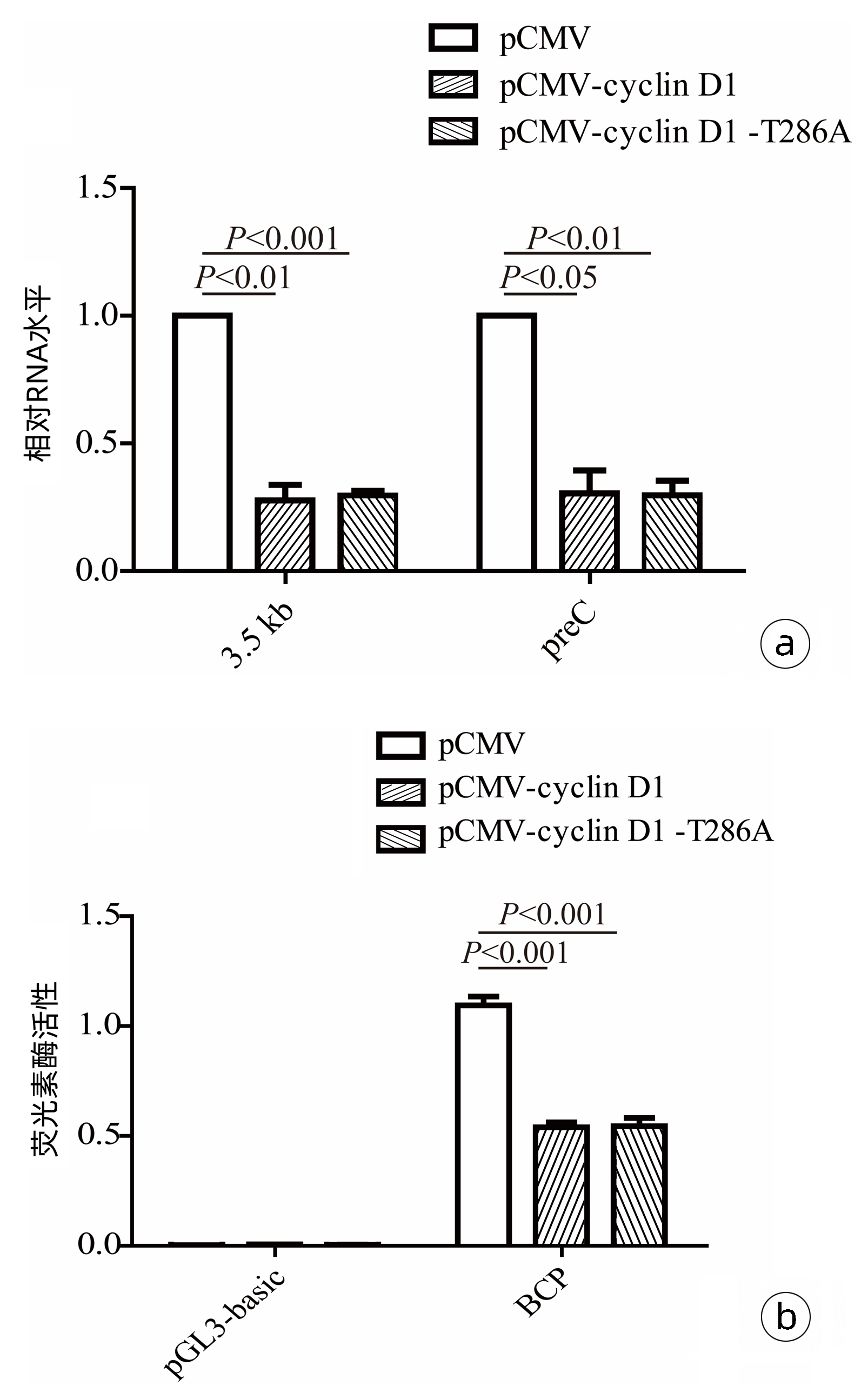
为了探究cyclin D1蛋白抑制HBV复制的可能环节,采用RT-qPCR方法,在HepG2细胞中检测了cyclin D1和cyclin D1(T286A)对HBV RNA水平的影响。结果显示,过表达cyclin D1和cyclin D1(T286A)均可显著抑制HepG2细胞中HBV的3.5 kb mRNA及preC mRNA水平(图 3a)。与此一致,双荧光素酶报告基因实验证实,过表达cyclin D1和cyclin D1(T286A)均显著抑制了HBV BCP启动子活性(图 3b)。
 图 3 外源表达cyclin D1对HBV转录水平的影响注:a,RT-qPCR检测cyclin D1和cyclin D1(T286A)过表达对HepG2细胞内HBV RNA的影响;b,双荧光素酶报告基因实验检测过表达cyclin D1和cyclin D1(T286A)对HBV BCP启动子活性的影响。Figure 3. Effects of exogenous expression of cyclin D1 on HBV transcript level
图 3 外源表达cyclin D1对HBV转录水平的影响注:a,RT-qPCR检测cyclin D1和cyclin D1(T286A)过表达对HepG2细胞内HBV RNA的影响;b,双荧光素酶报告基因实验检测过表达cyclin D1和cyclin D1(T286A)对HBV BCP启动子活性的影响。Figure 3. Effects of exogenous expression of cyclin D1 on HBV transcript level2.4 cyclin D1通过下调HNF1α和NTCP表达抑制HBV复制
为了进一步探究cyclin D1抑制HBV复制的可能机制,利用GSE83148数据集中122例CHB患者的肝脏转录组数据,分析了CCND1与10个已报道调控HBV复制的重要宿主基因的相关性(图 4a)。结果显示,CCND1水平与抑制HBV复制的APOBEC3G(r=0.575,P<0.001)、SMC5(r=0.341,P<0.001)和FOXM1(r=0.333,P<0.001)表达呈显著正相关,而与HBV进入受体NTCP(r=-0.511,P<0.001)和HBV复制正向调控转录因子HNF1α(r=-0.430,P<0.001)表达呈显著负相关。采用RT-qPCR方法,对与CCND1表达相关性较高的APOBEC3G、NTCP、HNF1α、SMC5和FOXM1进行了定量分析。结果显示,在HepG2细胞中过表达cyclin D1降低了HNF1α(P<0.001)和NTCP(P<0.01)的RNA水平,同时也抑制APOBEC3G和SMC5表达(图 4b)。
 图 4 CCND1与HBV调控基因的表达相关性分析注:a,CHB肝组织中CCND1与多个HBV调控基因的表达相关矩阵;b,RT-qPCR方法检测cyclin D1对HepG2细胞内APOBEC3G、NTCP、HNF1α、SMC5和FOXM1表达的影响。Figure 4. Expression correlation analysis of CCND1 and HBV-regulated genes in CHB liver tissues
图 4 CCND1与HBV调控基因的表达相关性分析注:a,CHB肝组织中CCND1与多个HBV调控基因的表达相关矩阵;b,RT-qPCR方法检测cyclin D1对HepG2细胞内APOBEC3G、NTCP、HNF1α、SMC5和FOXM1表达的影响。Figure 4. Expression correlation analysis of CCND1 and HBV-regulated genes in CHB liver tissues3. 讨论
病毒需要依附活细胞才能完成其生命周期,故病毒与宿主细胞之间存在大量互作调控。已有研究[4]报道,HBV复制依赖于细胞周期,肝细胞的快速增殖可以抑制HBV复制,但其机制尚未阐明。本研究发现细胞周期调控关键蛋白cyclin D1可以抑制HBV复制,机制上可能与cyclin D1抑制HNF1α和NTCP表达有关。
已有研究[4]发现,HBV感染能使原代肝细胞富集于G2/M期,同时促HBV复制的细胞转录因子如PPARA、RXRA和CEBPB等也在HBV感染时上调,表明HBV复制可能更易在增殖抑制的肝细胞中复制。在增殖旺盛的肝癌组织中,HBV cccDNA及HBV复制水平则显著低于癌旁组织,进一步证实增殖的肝细胞不利于病毒复制[13]。与上述发现一致,本研究通过对GEO数据库的肝脏转录组数据集分析发现,有7个细胞周期调控相关的基因表达与患者血清HBV DNA载量呈负相关,分别为MYC、TGFB1、CCND1、YWHAZ、HDAC1、YWHAB和E2F3。其中,已有文献报道MYC[14]及TGFB1[15]可以通过不同机制抑制HBV的复制,这与本研究的分析结果相符合。HDAC抑制剂[5]如FK228/SAHA以及敲减HDAC1基因,均可通过上调p21诱导细胞发生G0/G1期阻滞,进而促进HBV复制。但CCND1、YWHAZ、YWHAB和E2F3是否影响HBV复制目前未见报道。
CCND1基因编码的蛋白产物为cyclin D1。cyclin D1是调控细胞周期由G0/G1期向S期转换的关键因子。cyclin D1可与CDK4/6形成复合物,进入细胞核内使RB蛋白磷酸化,并使后者失去对转录因子E2F的抑制,进而上调cyclin E、cyclin A的表达,从而启动细胞向S期转换。cyclin D1主要经泛素-蛋白酶体系降解,其出核主要由出核运输蛋白CRM1介导[16],CRM1与cyclin D1结合依赖于cyclin D1第286位苏氨酸(T286)磷酸化[17]。当T286A突变导致该磷酸化基序丢失时,突变cyclin D1主要定位于细胞核内,可促进细胞异常增殖。本研究发现,表达外源cyclin D1及cyclin D1(T286A)突变体均能显著抑制HBsAg、HBeAg及HBV DNA水平,以及细胞内HBV RNA和HBV core蛋白水平,表明cyclin D1能抑制HBV复制。HBV BCP启动子主要调控3.5 kb mRNA的转录,其中pgRNA既可作为mRNA翻译合成病毒core蛋白和P蛋白,也可作为反转录的模板在核衣壳内合成rcDNA,而preC mRNA则主要翻译外泌的HBeAg。本研究发现cyclin D1能显著抑制BCP区的转录活性,提示cyclin D1可在转录水平抑制HBV复制。笔者团队也注意到,cyclin D1及cyclin D1(T286A)突变体对HBV复制的抑制作用未见明显不同,且二者对HepG2和Huh7细胞增殖的影响也不大,可能与本研究采用的HBV复制模型是增殖速度较快的肝癌细胞株有关,未来需要在原代肝细胞中加以验证。
目前认为,细胞周期影响HBV复制的机制主要有3个方面:(1)抑制HBV进入肝细胞的功能性受体NTCP表达,从而抑制HBV的从头感染[13, 18];(2)影响HBV复制调控分子的表达,如HNF4α、RXRA和CEBPB等[4],从而抑制HBV基因的转录;(3)影响HBV的成熟和释放[19-20]。本研究通过对数据分析发现,CHB患者肝组织中的CCND1与APOBEC3G、SMC5和FOXM1呈显著正相关,而与NTCP和HNF1α呈显著负相关。笔者团队的前期研究[13]发现,cyclin D1可抑制NTCP表达,提示cyclin D1也能抑制HBV对肝细胞的从头感染。HNF1α能够调控cccDNA的大部分调控元件(包括preS1启动子[21]、核心启动子[22]和增强子Ⅱ[23]),在HNF1α缺失的情况下HBV复制能力显著降低[24]。本研究进一步证实过表达cyclin D1能够抑制HNF1α和NTCP的表达,提示cyclin D1抑制HBV复制可能与其对HNF1α和NTCP的调控有关。
综上,本研究发现细胞周期关键蛋白cyclin D1可通过抑制HBV基因转录来抑制病毒复制,可为进一步阐明细胞周期调控HBV复制的分子机制提供理论依据,并为抗HBV新药研发提供潜在的分子靶点。
-

注:a,GSE84044数据集中与患者HBV DNA载量相关的肝组织宿主基因,样本按照HBV DNA载量的高低从左到右排列,蓝色代表低表达,红色代表高表达;b,765个与HBV DNA载量显著相关基因和116个细胞周期基因的交集;c,肝组织中CCND1水平与血清HBV DNA载量之间的相关性分析。
图 1 肝组织基因表达与血清HBV DNA的相关性分析
Figure 1. Correlation analysis of host gene expression in liver tissue and serum HBV DNA level

注:a、b,外源表达cyclin D1降低HepG2和Huh-7细胞HBV core蛋白的水平;c、d,外源表达cyclin D1下调HepG2和Huh-7细胞培养上清液中的HBsAg、HBeAg与HBV DNA水平。
图 2 外源表达cyclin D1降低HBV的复制水平
Figure 2. Exogenous expression of cyclin D1 reduces the replication level of HBV
注:a,RT-qPCR检测cyclin D1和cyclin D1(T286A)过表达对HepG2细胞内HBV RNA的影响;b,双荧光素酶报告基因实验检测过表达cyclin D1和cyclin D1(T286A)对HBV BCP启动子活性的影响。
图 3 外源表达cyclin D1对HBV转录水平的影响
Figure 3. Effects of exogenous expression of cyclin D1 on HBV transcript level
-
[1] GBD 2013 Mortality and Causes of Death Collaborators. Global, regional, and national age-sex specific all-cause and cause-specific mortality for 240 causes of death, 1990-2013: a systematic analysis for the Global Burden of Disease Study 2013[J]. Lancet, 2015, 385(9963): 117-171. DOI: 10.1016/S0140-6736(14)61682-2. [2] Chinese Society of Infectious Diseases, Chinese Medical Association; Chinese Society of Hepatology, Chinese Medical Association. Guidelines for the prevention and treatment of chronic hepatitis B (version 2019)[J]. J Clin Hepatol, 2019, 35(12): 2648-2669. DOI: 10.3969/j.issn.1001-5256.2019.12.007.中华医学会感染病学分会, 中华医学会肝病学分会. 慢性乙型肝炎防治指南(2019年版)[J]. 临床肝胆病杂志, 2019, 35(12): 2648-2669. DOI: 10.3969/j.issn.1001-5256.2019.12.007. [3] WANG J, HUANG H, LIU Y, et al. HBV genome and life cycle[J]. Adv Exp Med Biol, 2020, 1179: 17-37. DOI: 10.1007/978-981-13-9151-4_2. [4] XIA Y, CHENG X, LI Y, et al. Hepatitis B virus deregulates the cell cycle to promote viral replication and a premalignant phenotype[J]. J Virol, 2018, 92(19): e00722-18. DOI: 10.1128/JVI.00722-18. [5] YANG Y, YAN Y, CHEN Z, et al. Histone deacetylase inhibitors romidepsin and vorinostat promote hepatitis B virus replication by inducing cell cycle arrest[J]. J Clin Transl Hepatol, 2021, 9(2): 160-168. DOI: 10.14218/JCTH.2020.00105. [6] OTTO T, SICINSKI P. Cell cycle proteins as promising targets in cancer therapy[J]. Nat Rev Cancer, 2017, 17(2): 93-115. DOI: 10.1038/nrc.2016.138. [7] TCHAKARSKA G, SOLA B. The double dealing of cyclin D1[J]. Cell Cycle, 2020, 19(2): 163-178. DOI: 10.1080/15384101.2019.1706903. [8] QI Z, LI G, HU H, et al. Recombinant covalently closed circular hepatitis B virus DNA induces prolonged viral persistence in immunocompetent mice[J]. J Virol, 2014, 88(14): 8045-8056. DOI: 10.1128/JVI.01024-14. [9] WANG M, GONG Q, ZHANG J, et al. Characterization of gene expression profiles in HBV-related liver fibrosis patients and identification of ITGBL1 as a key regulator of fibrogenesis[J]. Sci Rep, 2017, 7: 43446. DOI: 10.1038/srep43446. [10] ZHOU W, MA Y, ZHANG J, et al. Predictive model for inflammation grades of chronic hepatitis B: Large-scale analysis of clinical parameters and gene expressions[J]. Liver Int, 2017, 37(11): 1632-1641. DOI: 10.1111/liv.13427. [11] RITCHIE ME, PHIPSON B, WU D, et al. limma powers differential expression analyses for RNA-sequencing and microarray studies[J]. Nucleic Acids Res, 2015, 43(7): e47. DOI: 10.1093/nar/gkv007. [12] R Core Team (2022). R: A language and environment for statistical computing[EB/OL]. https://www.R-project.org/ .[13] KANG J, WANG J, CHENG J, et al. Down-regulation of NTCP expression by cyclin D1 in hepatitis B virus-related hepatocellular carcinoma has clinical significance[J]. Oncotarget, 2017, 8(34): 56041-56050. DOI: 10.18632/oncotarget.10241. [14] JUNG YJ, KIM JW, PARK SJ, et al. c-Myc-mediated overexpression of miR-17-92 suppresses replication of hepatitis B virus in human hepatoma cells[J]. J Med Virol, 2013, 85(6): 969-978. DOI: 10.1002/jmv.23534. [15] ALMAJHDI FN, AL-QUDARI AY, HUSSAIN Z. Differential expression of transforming growth factor-β1 and HBx enhances hepatitis B virus replication and augments host immune cytokines and chemokines[J]. Ann Hepatol, 2013, 12(3): 408-415. [16] ALT JR, CLEVELAND JL, HANNINK M, et al. Phosphorylation-dependent regulation of cyclin D1 nuclear export and cyclin D1-dependent cellular transformation[J]. Genes Dev, 2000, 14(24): 3102-3114. DOI: 10.1101/gad.854900. [17] DIEHL JA, CHENG M, ROUSSEL MF, et al. Glycogen synthase kinase-3beta regulates cyclin D1 proteolysis and subcellular localization[J]. Genes Dev, 1998, 12(22): 3499-3511. DOI: 10.1101/gad.12.22.3499. [18] YAN Y, ALLWEISS L, YANG D, et al. Down-regulation of cell membrane localized NTCP expression in proliferating hepatocytes prevents hepatitis B virus infection[J]. Emerg Microbes Infect, 2019, 8(1): 879-894. DOI: 10.1080/22221751.2019.1625728. [19] KÖNIG A, YANG J, JO E, et al. Efficient long-term amplification of hepatitis B virus isolates after infection of slow proliferating HepG2-NTCP cells[J]. J Hepatol, 2019, 71(2): 289-300. DOI: 10.1016/j.jhep.2019.04.010. [20] ELLER C, HEYDMANN L, COLPITTS CC, et al. A genome-wide gain-of-function screen identifies CDKN2C as a HBV host factor[J]. Nat Commun, 2020, 11(1): 2707. DOI: 10.1038/s41467-020-16517-w. [21] RANEY AK, EASTON AJ, MILICH DR, et al. Promoter-specific transactivation of hepatitis B virus transcription by a glutamine- and proline-rich domain of hepatocyte nuclear factor 1[J]. J Virol, 1991, 65(11): 5774-5781. DOI: 10.1128/JVI.65.11.5774-5781.1991. [22] ZHENG Y, LI J, OU JH. Regulation of hepatitis B virus core promoter by transcription factors HNF1 and HNF4 and the viral X protein[J]. J Virol, 2004, 78(13): 6908-6914. DOI: 10.1128/JVI.78.13.6908-6914.2004. [23] WANG WX, LI M, WU X, et al. HNF1 is critical for the liver-specific function of HBV enhancer Ⅱ[J]. Res Virol, 1998, 149(2): 99-108. DOI: 10.1016/s0923-2516(98)80085-x. [24] QUASDORFF M, HÖSEL M, ODENTHAL M, et al. A concerted action of HNF4alpha and HNF1alpha links hepatitis B virus replication to hepatocyte differentiation[J]. Cell Microbiol, 2008, 10(7): 1478-1490. DOI: 10.1111/j.1462-5822.2008.01141.x. 期刊类型引用(3)
1. 张晓威,王楠,代梦格,刘瑞娟,马亭. 白花蛇舌草-半枝莲药对治疗胃癌的物质基础和作用机制研究进展. 中国临床药理学与治疗学. 2024(07): 831-840 .  百度学术
百度学术2. 周召,鲁凤民. 细胞核受体结合蛋白1通过促进乙肝病毒核心启动子活性增强病毒复制. 国际病毒学杂志. 2024(05): 371-375 . 百度学术3. 何蓉,何小清,李君怡,易静. 高危型HPV感染与宫颈病变组织中cyClinD1、Ki-67和HER-2蛋白表达的关联. 中华医院感染学杂志. 2024(24): 3793-3797 . 百度学术其他类型引用(0)
-


 下载:
下载:


 下载:
下载:

 百度学术
百度学术

