
 PDF下载 ( 48190 KB)
PDF下载 ( 48190 KB)

克老素衍生肽7对慢性胰腺炎小鼠模型胰腺纤维化的影响及其机制
DOI: 10.12449/JCH260419
Effect of Klotho-derived peptide 7 on pancreatic fibrosis in a mouse model of chronic pancreatitis and its mechanism
-
摘要:
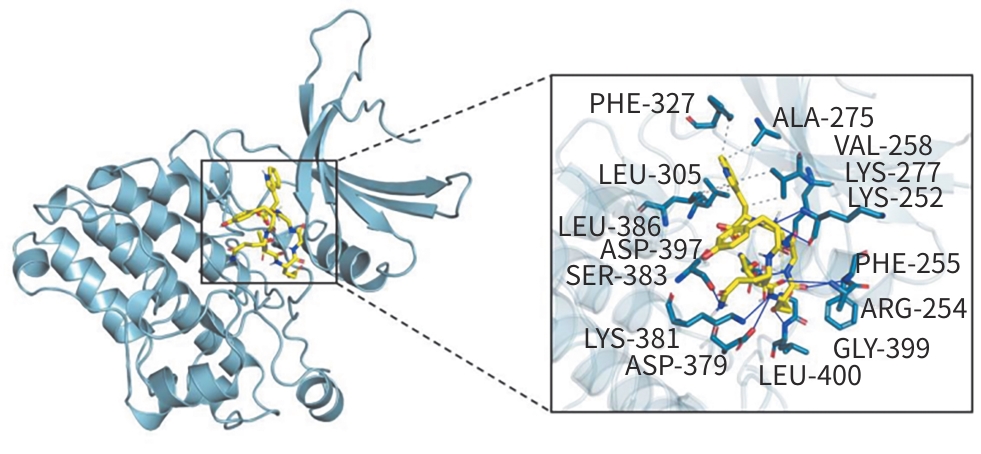
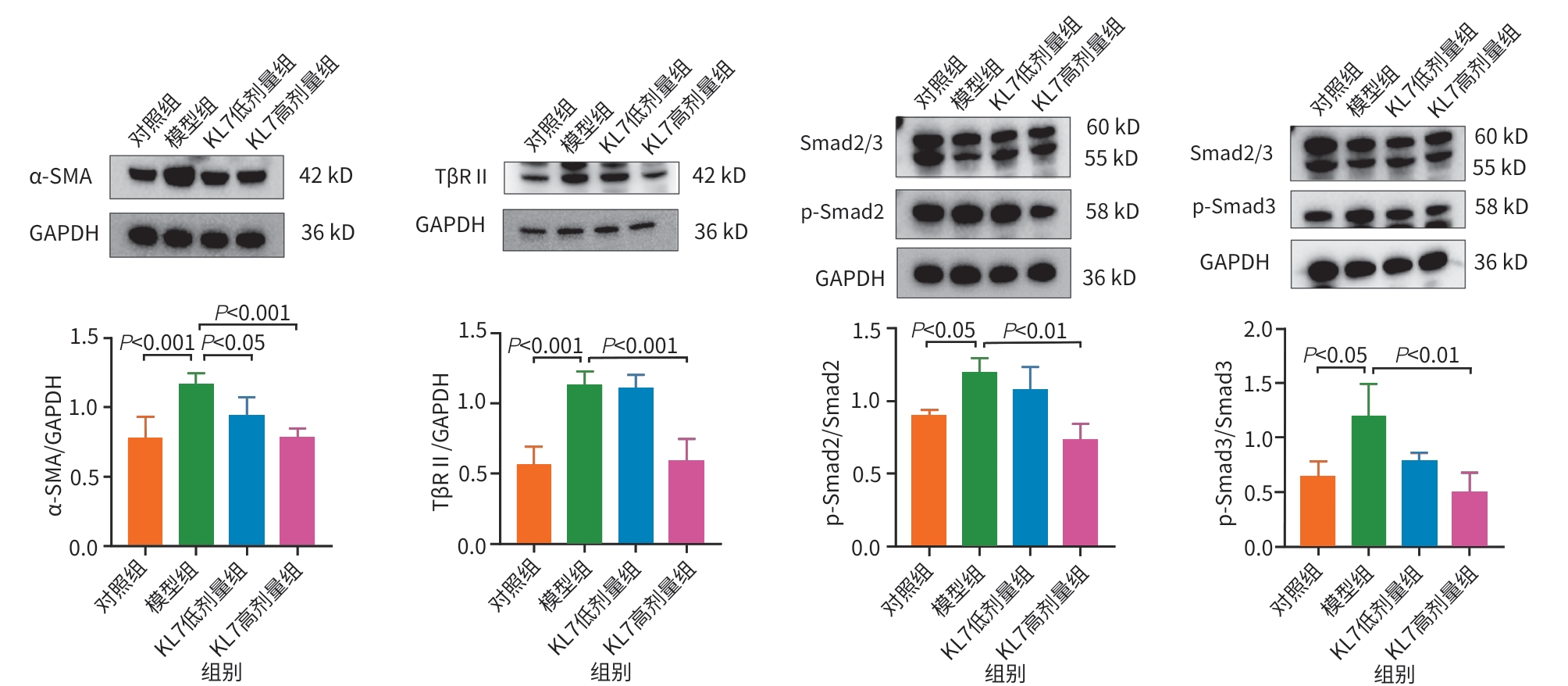
目的 通过观察克老素衍生肽7(KL7)对雨蛙素诱导的慢性胰腺炎(CP)小鼠模型的影响,了解该药物抗胰腺纤维化的作用机制,进而为临床用药提供依据。 方法 将40只雄性BALB/c小鼠随机分为对照组、模型组、KL7低剂量组(2 mg/kg)和KL7高剂量组(4 mg/kg),每组10只。除对照组外,其余各组小鼠腹腔注射雨蛙素(50 μg/kg),每天注射6次,每次间隔1 h,每周注射3 d,连续注射4周,进行CP造模。KL7低剂量、KL7高剂量组分别给予不同剂量的KL7治疗,每天给药1次,连续4周。活体成像观察KL7在胰腺中的富集情况;分子对接检测KL7与转化生长因子β Ⅱ型受体(TβRⅡ)的结合情况;称量小鼠体重及胰腺湿重;苏木素-伊红染色观察胰腺组织的病理变化;马松染色观察胰腺组织纤维化程度;免疫组化染色检测α-平滑肌肌动蛋白(α-SMA)、Ⅰ型胶原α1链(COL1A1)的表达情况;蛋白质印迹法检测胰腺组织中α-SMA、TβRⅡ、磷酸化的母系抗十五表态蛋白同源物2/3(p-Smad2/3)蛋白的表达水平。计量资料多组间比较采用单因素方差分析,进一步两两比较采用LSD-t和Dunnett’s-T3检验。 结果 KL7能够显著富集在CP小鼠胰腺组织,与TβRⅡ蛋白存在强结合活性。与对照组相比,模型组小鼠胰腺湿量和相对胰腺质量均显著降低(P<0.000 1),胰腺组织结构紊乱、炎细胞浸润增加,纤维化程度显著增加,α-SMA和COL1A1阳性面积均显著增加(P值均<0.000 1),α-SMA、TβRⅡ和p-Smad2/3蛋白表达水平显著升高(P值均<0.05)。与模型组相比,KL7高剂量组小鼠胰腺湿量和相对胰腺质量均显著升高(P值均<0.01),胰腺组织结构损伤和炎细胞浸润得到缓解,纤维化程度显著降低,α-SMA和COL1A1阳性面积均显著降低(P值均<0.001),α-SMA、TβRⅡ和p-Smad2/3蛋白表达水平均显著降低(P值均<0.01)。 结论 KL7对CP小鼠胰腺纤维化具有显著的靶向治疗效果,其作用机制涉及KL7与TβRⅡ特异性结合,从而抑制TGF-β/Smad信号通路的激活。 Abstract:Objective To investigate the anti‑pancreatic fibrosis mechanism of Klotho‑derived peptide 7 (KL7) by observing its effect on a mouse model of chronic pancreatitis (CP) induced by cerulean, and to provide a basis for clinical medication. Methods A total of 40 male BALB/c mice were randomly divided into control group, model group, low-dose KL7 group (2 mg/kg), and high-dose KL7 group (4 mg/kg), with 10 mice in each group. All mice except those in the control group were given intraperitoneal injection of cerulean (50 μg/kg) 6 times a day at an interval of 1 hour, twice a week for 4 consecutive weeks to establish a model of CP. The mice in the low-dose KL7 group and the high-dose KL7 group were treated with different doses of KL7 once a day for 4 consecutive weeks. In vivo imaging was used to observe the accumulation of KL7 in the pancreas; molecular docking was used to detect the binding of KL7 to transforming growth factor-β type Ⅱ receptor (TβRⅡ); the mice were measured in terms of body weight and pancreatic weight; HE staining was used to observe the pathological changes of pancreatic tissue; Masson staining was used to observe the degree of pancreatic fibrosis; immunohistochemical staining was used to measure the expression of α-smooth muscle actin (α-SMA) and type Ⅰ collagen (COL1A1); Western blotting was used to measure the protein expression levels of α-SMA, TβRII, and phosphorylated small mothers against decapentaplegic homolog 2/3 (p-Smad2/3) in pancreatic tissue. A one-way analysis of variance was used for comparison of continuous data between multiple groups, and the least significant difference t-test and the Dunnett’s-T3 test were used for further comparison between two groups. Results KL7 was significantly enriched in the pancreatic tissue of CP mice, and there was a strong binding activity between KL7 and TβRⅡ. Compared with the control group, the model group had significant reductions in pancreatic mass and relative pancreatic mass (P<0.000 1), with disordered structure of pancreatic tissue, an increase in inflammatory cell infiltration, and significant increases in fibrosis degree, the positive areas of α-SMA and COL1A1 (P<0.000 1), and the protein expression levels of α-SMA, TβRⅡ, and p-Smad2/3 (P<0.05). Compared with the model group, the high-dose KL7 group had significant increases in pancreatic mass and relative pancreatic mass (P<0.01), with alleviation of structural damage of pancreatic tissue and inflammatory cell infiltration, a significant reduction in fibrosis degree, and significant reductions in the positive areas of α-SMA and COL1A1 (P<0.001) and the protein expression levels of α-SMA, TβRⅡ, and p-Smad2/3 (P<0.01). Conclusion KL7 has a significant targeted therapeutic effect on pancreatic fibrosis in CP mice through specific binding of KL7 to TβRⅡ, thereby inhibiting the activation of the TGF-β/Smad signaling pathway. -
Key words:
- Pancreatitis, Chronic /
- Fibrosis /
- Klotho-Derived Peptide
-
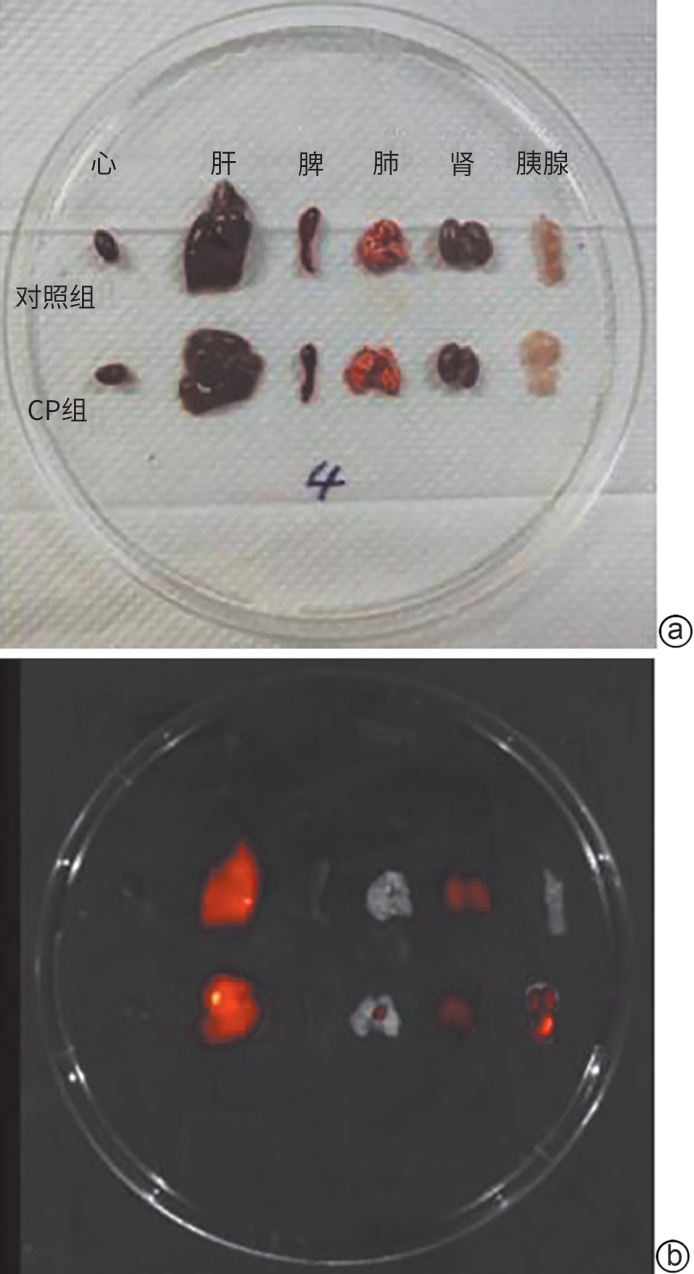
注: a,脏器明场;b,脏器克老素衍生肽7-花青素7荧光成像。
图 1 克老素衍生肽7在各组小鼠主要脏器中的富集情况
Figure 1. Enrichment of KL7 in the main organs of mice in various group
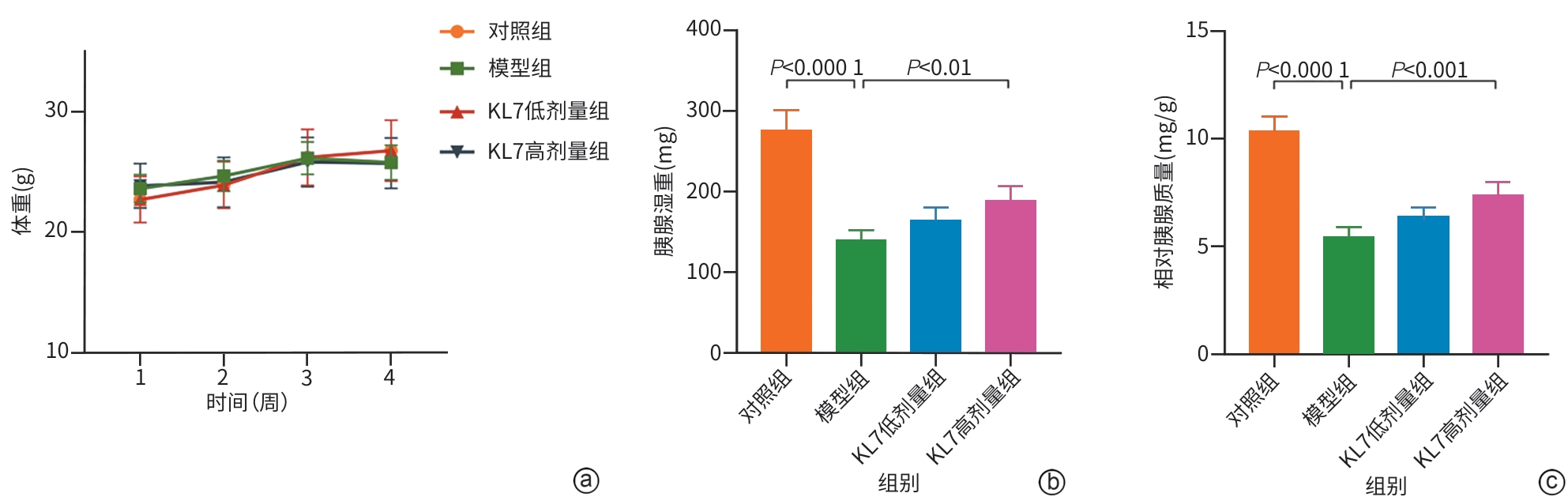
注: a,各组小鼠体重;b,各组小鼠胰腺湿重;c,各组小鼠相对胰腺质量。KL7,克老素衍生肽7。
图 3 各组小鼠体重及胰腺湿重情况
Figure 3. Body weight and pancreatic weight of mice in various group

注: KL7,克老素衍生肽7。
图 4 各组小鼠胰腺组织大体形态和苏木精-伊红染色结果(×100)
Figure 4. Gross morphology and HE staining results of pancreatic tissue in various group

注: a,马松染色(×100);b,α-SMA表达情况(免疫组织化学染色,×100);c,COL1A1表达情况(免疫组织化学染色,×100);d, 各组小鼠胰腺组织马松染色和免疫组织化学染色面积半定量结果。α-SMA,α平滑肌肌动蛋白;COL1A1,Ⅰ型胶原α1链;KL7,克老素衍生肽7。
图 5 各组小鼠胰腺组织染色情况
Figure 5. Staining status of pancreatic tissues in various groups of mice
-
[1] HINES OJ, PANDOL SJ. Management of chronic pancreatitis[J]. BMJ, 2024, 384: e070920. DOI: 10.1136/bmj-2023-070920. [2] SINGH VK, YADAV D, GARG PK. Diagnosis and management of chronic pancreatitis: A review[J]. JAMA, 2019, 322( 24): 2422- 2434. DOI: 10.1001/jama.2019.19411. [3] CAI QY, TAN K, ZHANG XL, et al. Incidence, prevalence, and comorbidities of chronic pancreatitis: A 7-year population-based study[J]. World J Gastroenterol, 2023, 29( 30): 4671- 4684. DOI: 10.3748/wjg.v29.i30.4671. [4] de THIERENS N, VERDONK RC, LÖHR JM, et al. Chronic pancreatitis[J]. Lancet, 2025, 404( 10471): 2605- 2618. DOI: 10.1016/S0140-6736(24)02187-1. [5] KALE A, SANKRITYAYAN H, ANDERS HJ, et al. Epigenetic and non-epigenetic regulation of Klotho in kidney disease[J]. Life Sci, 2021, 264: 118644. DOI: 10.1016/j.lfs.2020.118644. [6] ROSTAMZADEH F, MOOSAVI-SAEED Y, YEGANEH-HAJAHMADI M. Interaction of klotho and sirtuins[J]. Exp Gerontol, 2023, 182: 112306. DOI: 10.1016/j.exger.2023.112306. [7] PRUD’HOMME GJ, KURT M, WANG QH. Pathobiology of the klotho antiaging protein and therapeutic considerations[J]. Front Aging, 2022, 3: 931331. DOI: 10.3389/fragi.2022.931331. [8] YUAN Q, REN Q, LI L, et al. A Klotho-derived peptide protects against kidney fibrosis by targeting TGF-β signaling[J]. Nat Commun, 2022, 13: 438. DOI: 10.1038/s41467-022-28096-z. [9] CHEN XW, TAN HS, XU J, et al. Klotho-derived peptide 6 ameliorates diabetic kidney disease by targeting Wnt/β-catenin signaling[J]. Kidney Int, 2022, 102( 3): 506- 520. DOI: 10.1016/j.kint.2022.04.028. [10] SHEN JL, BIN WY, LIN XN, et al. Klotho protein: A multifaceted guardian of healthy aging and its therapeutic potential[J]. Int J Nanomedicine, 2025, 20: 7251- 7270. DOI: 10.2147/IJN.S514516. [11] ZHAO XY, HAN DH, ZHAO C, et al. New insights into the role of Klotho in inflammation and fibrosis: Molecular and cellular mechanisms[J]. Front Immunol, 2024, 15: 1454142. DOI: 10.3389/fimmu.2024.1454142. [12] PRUD’HOMME GJ, GLINKA Y, KURT M, et al. Systemic Klotho therapy protects against insulitis and enhances beta-cell mass in NOD mice[J]. Biochem Biophys Res Commun, 2020, 525( 3): 693- 698. DOI: 10.1016/j.bbrc.2020.02.123. [13] ARBEL RUBINSTEIN T, REUVENI I, HESIN A, et al. A transgenic model reveals the role of klotho in pancreatic cancer development and paves the way for new klotho-based therapy[J]. Cancers, 2021, 13( 24): 6297. DOI: 10.3390/cancers13246297. [14] APTE MV, PIROLA RC, WILSON JS. Pancreatic stellate cells: A starring role in normal and diseased pancreas[J]. Front Physiol, 2012, 3: 344. DOI: 10.3389/fphys.2012.00344. [15] BYNIGERI RR, JAKKAMPUDI A, JANGALA R, et al. Pancreatic stellate cell: Pandora's box for pancreatic disease biology[J]. World J Gastroenterol, 2017, 23( 3): 382- 405. DOI: 10.3748/wjg.v23.i3.382. [16] KONG FY, PAN YY, WU D. Activation and regulation of pancreatic stellate cells in chronic pancreatic fibrosis: A potential therapeutic approach for chronic pancreatitis[J]. Biomedicines, 2024, 12( 1): 108. DOI: 10.3390/biomedicines12010108. [17] MASAMUNE A, WATANABE T, KIKUTA K, et al. Roles of pancreatic stellate cells in pancreatic inflammation and fibrosis[J]. Clin Gastroenterol Hepatol, 2009, 7( 11 Suppl): S48- S54. DOI: 10.1016/j.cgh.2009.07.038. [18] ZHENG MF, LI HY, GAO YH, et al. Vitamin D3 analogue calcipotriol inhibits the profibrotic effects of transforming growth factor- β1 on pancreatic stellate cells[J]. Eur J Pharmacol, 2023, 957: 176000. DOI: 10.1016/j.ejphar.2023.176000. [19] CHUNG JY, CHAN MK, LI JS, et al. TGF-β signaling: From tissue fibrosis to tumor microenvironment[J]. Int J Mol Sci, 2021, 22( 14): 7575. DOI: 10.3390/ijms22147575. -

 下载:
下载:
 本文二维码
本文二维码
计量
- 文章访问数: 164
- HTML全文浏览量: 61
- PDF下载量: 59
- 被引次数: 0

